Proposal to Import Bombus terrestris (Bt) onto Mainland Australia for Crop Pollination Purposes
Factual Errors, Misleading Statements and Half Truths in this Australian Hydroponic and Greenhouse Association Report
|
|
AHGA Report Factual Errors, Misleading Statements or Half Truths
|
|
The Actual Facts
|
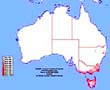
Potential Distribution of Feral Bumblebees
|
AHGA report page iii
The McClay CLIMEX study indicates the likelihood of only limited distribution of B. terrestris (subspecies audax) on mainland Australia should it arrive either by accident or design. The study predicts these areas will be restricted to the wetter areas of Victoria, the south-west corner of Western Australia, and a limited area of NSW across the north-eastern border of Victoria...
|
|
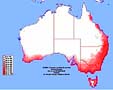
The real range of feral bumblebees in Australia could be much larger!
The McClay CLIMEX study actually produced two different predicted distribution maps. Dr McClay stated (see AHGA report, page 95) that the smaller distribution 'represents an absolute minimum predicted range for this subspecies in Australia... it should be expected that even this subspecies could establish in broader areas of Australia, possibly approaching [this distribution]':
The AHGA rejects Dr McClay's larger distribution on the basis of research by Dr Madeleine Beekman; but Dr Beekman says the AHGA is misinterpreting her results. (More details.)
|
|
The Native Bee Alternative: Blue Banded Bees
|
|
AHGA Report Says
|
|
The Actual Facts
|
AHGA report page xix
Research on Amegilla spp. [blue banded bees] has been conducted for several years now through the University of Western Sydney and the University of Adelaide, but to date there have been no scientific publications... |
|
A scientific paper on Melissa Bell's University of Western Sydney research on Amegilla blue banded bees and tomatoes has just been published in Journal of Economic Entomology (Vol 99: 437-442), and reviewed in Aussie Bee Online Article 9. Melissa found that blue banded bees foraged successfully in the greenhouse, nested in artificial nest blocks and pollinated the tomatoes as efficiently as the vibrating wand tools currently used by tomato growers.
Another scientific paper on Dr Katja Hogendoorn's University of Adelaide research on Amegilla blue banded bees and tomatoes has now appeared in the same high quality, peer reviewed journal in June 2006: Journal of Economic Entomology (Vol 99: 828-833). Katja found that the plants pollinated by blue banded bees produced tomatoes 15-20% heavier than those pollinated by the currently used vibrating wand tool. |
AHGA report page xix
Discovering how to break diapause [winter dormant period] without negative effects will be essential for all-year round use in greenhouses. However, no information on conditions for inducing or terminating diapause has been published, only that it seems to occur in response to low temperatures. |
|
Dr Katja Hogendoorn says her 2.5 years of research so far have delivered protocols for year round breeding of blue banded bees in large numbers. Methods for breaking winter dormancy are in place but need fine tuning.
It is important to note that the exact methods for breeding bumblebees in industrial quantities have also never been published, for commercial reasons. The same confidentiality applies to details of the breeding of blue banded bees. This information is currently being transferred to one of the industry partners in the blue banded bee project (Biological Services).
|
AHGA report page xix
Nor is information available on the temperature range under which these bees will provide efficient pollination.... |
|
Dr Katja Hogendoorn says blue banded bees actively forage between temperatures of 20 and 40 degrees C (some as low as 18 degrees C), which is well within the day-time range used in commercial Australian greenhouses. Temperatures in the 30s, which regularly occur many Australian greenhouses, do not depress blue banded bee performance. In contrast, temperatures in the 30s markedly reduce bumblebee performance! (More details) |
AHGA report page xix
Information gleaned from 'The Buzz', a newsletter put out by the researchers, and by an article in Practical Hydroponics and Greenhouses (Sedgley et al. 2004) suggests that a practical solution is not on the horizon. |
|
The fact that the main breeding program is currently being moved to one of the project's industry partners, Biological Services, clearly indicates that the blue banded bee researchers are well on their way towards achieving a practical solution for the greenhouse tomato industry.
Subsequent updates by the Adelaide blue banded bee research group, published in Practical Hydroponics and Greenhouses magazine in 2004 and 2005, listed progress made in solving key technical problems such as breaking diapause and mass breeding.
A recent update on the blue banded bee research has also been published in the May/June 2006 issue of Practical Hydroponics and Greenhouses magazine (Issue 88, pp 24-26).
A statement by Dr Katja Hogendoorn in May 2006 concludes, 'Good progress has been made. Depending on the financial support for the project, the [remaining] issues can be sorted out in three years time. After that, a breeding facility needs to be built, and breeding stock will need to be built up for a year. Altogether, six years seems to be a realistic estimate.'
|
AHGA report page xix
Further difficulties stem from the fact that screening is necessary to keep them [blue banded bees] within the greenhouse... |
|
Why is this such a problem? In order to prevent exotic bumblebees from escaping into the Australian bushland, the AHGA proposes that all tomato growers wishing to use bumblebees must also fully screen their greenhouses: 'Secure greenhouses would be those suitably screened at all vents to prevent escape of any bumblebees, and to have an annex at all entry points with positive air pressure.' (see AHGA report page 65)
|
AHGA report page xx
Hogendoorn (2004) envisages growers rearing brood on the greenhouse site, as it is difficult to provide pollen artificially. |
|
The AHGA needs to update their information! This statement was made just three months after the start of the blue banded bee research, back in 2004.
Dr Katja Hogendoorn has now developed methods for breeding blue banded bees, in large quantities, year round, in a specialised breeding facility. Commercial tomato growers will not need to breed their own blue banded bees. (More details)
|
AHGA report page xx
Proponents of developing a native bee such as Amegilla as a commercial pollinator should also bear in mind that shipping a single species around Australia, without regard to whether it is native in all those regions, or whether there are strain differences within the same species, raises the same issues as whether Bombus impatiens should be used in western North America when it is native only to the east of the continent, or B. terrestris should be moved around Europe without regard to its several subspecies. |
|
Some blue banded bee species are very widespread and common in Australia. Important research is already in progress identifying the different species and strains of blue banded bees and their distributions. A species that is native to wide areas of Australia will be chosen for mass production. |
AHGA report page xx
Currently, the [blue banded bee] research work being conducted on the mainland appears to be seriously under-funded... |
|
The Australian greenhouse tomato industry 'would benefit from an annual cash bonus of over $8 million' (AHGA report page xviii) from the acquisition of an efficient pollinating bee for their crops. Why does the AHGA not fund the blue banded bee research program instead of criticising it for being under-funded? |
|
|
|
|
AHGA Report Says
|
|
The Actual Facts
|
AHGA report page 12
Visitation records [of bumblebees in New Zealand] show marked preferences for introduced rather than native plants, and they are rare in native vegetation. Populations of native bees are competing successfully...
and AHGA report page 27
.....the situation reported from New Zealand, where introduced plants were strongly preferred [by B. terrestris bumblebees] (Donovan 1980; Donovan and Macfarlane 1984).
|
|
New Zealand's ecology is very different from Australia's so their experience is not a valid predictor of the likely impact of bumblebees in Australia.
Dr Andrew Hingston points out that 91% of New Zealand's plants are now introduced species so it is hardly surprising that bumblebees appear to 'prefer' introduced plants. It is true that B. terrestris has been recorded foraging on more species of introduced plants (at least 400; Donovan and Macfarlane 1984) than native plants (at least 53; Macfarlane and Gurr 1995) in New Zealand. However, this cannot be interpreted as preference because there are ten times as many species of introduced plants (20,551) than native plants (2,055) in New Zealand (Williams and West 2000). Hence, B. terrestris has been recorded foraging on 1.95% of introduced, and 2.58% of native, plant species growing in New Zealand!
Furthermore, New Zealand possesses only 23 species of native bees compared with over 1,500 native bee species in Australia. So we have many more vulnerable native bee species in Australia that could be affected by competition with bumblebees.
|
|
Bumblebee Impact on Native Bees
|
|
AHGA Report Says
|
|
The Actual Facts
|
AHGA report p29
Two independent analyses of the statistics employed in [Hingston and McQuillan's 1999] study (Anon 2004, A. McClay 2005b) were conducted. Both stated that because of a design flaw, the study does not have sufficient statistical power to confirm a displacement effect.
|
|
Scientific papers are considered to carry little weight unless they are published in refereed scientific journals. Hingston and McQuillan published their paper in a refereed scientific journal seven years ago, giving ample time for any flaws to be exposed by subsequent refereed scientific papers. However, Dr Hingston is not aware of any such refereed papers and the AHGA does not list any in this report.
Instead the AHGA report quotes two references, which do not appear to have been refereed, as evidence of criticism of this paper. The first of these 'Anon, 2004' is anonymous, and is not included in their reference list. The second (McClay 2005) is a private analysis prepared for the AHGA, which they have not reproduced as part of their report. These unsubstantiated documents can hardly be taken as serious proof of a 'design flaw' in the Hingston and McQuillan paper.
|
| AHGA report pages 29-30
Goulson et al 2002 'do not suggest that bumble bees are having a significant impact upon native bee communities... the addition of (bumblebees) is perhaps unlikely to have an appreciable effect on small, short-tongued native bees'
|
|
Dr Andrew Hingston points out that this study by Goulson, Stout and Kells only ran from 15 November to 10 December, a time of year when Tasmanian bumblebee populations remain relatively small (Hingston and McQuillan 1998a) and Exoneura and Halictinae native bees are dominant (Hingston 1998; Goulson et al. 2002). Exoneura and most Halictinae have broad foraging preferences (Armstrong 1979; Hingston 1999) so they may be able to withstand some bumblebee competition by foraging from plants that are not heavily exploited by bumblebees.
In contrast, many of the Tasmanian Colletidae short-tongued native bees are most abundant after late December (Hingston 1998) when bumblebee populations are largest. So the Colletidae bees are the family of Tasmanian bees with foraging profiles most similar to bumblebees (Hingston and McQuillan 1998a). About half of the described species of native bees in Australia are in Colletidae (Cardale 1993). Many of these native bees are highly vulnerable to competition because they have very narrow foraging profiles (Armstrong 1979; Cardale 1993; Hingston 1999).
|
| AHGA report page 30
It may thus be incorrect to state that B. terrestris will provide an add-on effect to that of honeybees by competing with native bees at hours when honeybees do not normally forage.
|
|
Dr Andrew Hingston has studied patterns of nectar consumption during the day by bumblebees and Apis honeybees from Eucalyptus ovata in Tasmania. The results clearly show that bumblebees can reduce nectar availability at times when it is too cold for honeybees to forage, such as early mornings and throughout cold days. Dr Hingston is about to submit a paper on this research to a scientific journal. |
AHGA report page 57
It is difficult to see that any case for significant competition of bumblebees with native bees and birds could be sustained.
|
|
A study near Hobart showed that bumblebees were foraging on at least 66 species of native plants from 21 families (Hingston and McQuillan 1998a). These plants provided at least part of the food resources for all the subgenera of Tasmanian native bees and species of nectar-feeding birds seen in the study. This shows that many species of native bees and birds are at risk of competition from bumblebees.
Although it is notoriously difficult to prove competition effects in bees, it is very likely that bumblebees would compete with many Australian native bee species. Here are some of the reasons (see Goulson 2003 for more details):
Most of Australia's 1500 native bee species:
--- are solitary (just one female builds each nest)
--- store no food resources in their tiny nests and must individually gather all the nectar and pollen they need to provision their brood cells day by day
--- must often complete their nests within a few weeks
--- cannot fly in inclement weather
--- can often fly only a few hundred metres from their nests
--- must discover good food sources for themselves by trial and error
In contrast, bumblebees:
--- are social (building nests with up to 500 bees)
--- store some food in their nests
--- can nest throughout the spring and summer
--- can fly in cooler overcast weather and can gather large quantities of nectar and pollen from flowers before the solitary native bees begin flying
--- can fly at least 4 km from their nests
--- can learn from nest mates about the discovery and scent of particularly good food sources
Goulson (2003) concludes, 'It is hard to conceive how the introduction of these exotic species [honeybees and bumblebees] ... could not have substantially altered the diversity and abundance of native bees.'
|
|
Bumblebee Impact on Native Plants
|
|
AHGA Report Says
|
|
The Actual Facts
|
| AHGA report page 31
[Re: Hingston, 2005 study of bumblebee preferences for introduced or native plants] Hingston shows a complete disregard for the wealth of published information on the intricacies of foraging behaviour in bumblebees (see Prys-Jones and Corbet (1991) and Goulson (2003a) for a partial review of this well-researched subject).
|
|
Dr Hingston's paper (Hingston 2005a, page 31) actually reports several lines from one of the references which the AHGA claims was completely disregarded: 'It is also consistent with Prys-Jones and Corbet (1991), who noted that almost all species of British bumblebees, including B. terrestris, readily forage in parts of Britain where the native vegetation has been replaced by plants from all over the world.'
This reference supports Dr Hingston's findings by stating that bumblebees do not show preferences among flowers on the basis of geographic origin.
|
| AHGA report page 33
[Re: Hingston and McQuillan, 1998 study of nectar robbing in bumblebees] No bees were apparently actually observed biting the holes, despite the 13 days of observation.
|
|
This AHGA statement is incorrect. The Results section, and Table 1 in particular, of Hingston and McQuillan (1998b) clearly show that bumblebees were observed biting holes in the corollas of E. impressa. |
| AHGA report page 34
[Re: Discussion in Hingston and McQuillan 1998a of bumblebee impact on commercial honey production] Thus the likelihood of B. terrestris establishing in any numbers in west coast old growth forests, and then competing for leatherwood nectar with managed hives of honeybees, is extremely unlikely.
|
|
Frequent observations of bumblebees in large numbers in native vegetation in western Tasmania, including many observations of them feeding on flowers of leatherwood (Hingston et al. 2002; Hingston 2006), indicate that bumblebees are a potential competitor with commercial honeybees for this resource. Because the seasonal peak in bumblebee abundance coincides with the flowering of leatherwood, this impact could be serious.
The potential for bumblebees to harm the Tasmanian honey industry is highlighted by the observations of Stout et al. (2002, page 432) 'In Tasmania, B. terrestris may be competitively excluding A. mellifera from L. arboreus, C. scoparius and other species (e.g. Digitalis purpurea L., Scrophulariaceae). A. mellifera only forages from these species in large numbers when B. terrestris is absent.'
Indeed, the Tasmanian honey industry now appears to be very concerned about the threat that competition from bumblebees poses to their industry. Dr Hingston was invited to present a seminar at the Tasmanian Beekeepers' Association AGM last year, and delegates were very worried about this threat. Dr Hingston said that beekeepers did not seem very interested in having more research done into the impacts of bumblebees; they just wanted to know how to get rid of them. |
AHGA report page 56
...no significant effect on native plants on the mainland is anticipated.
|
|
For effective pollination, the shape of the flower must match the shape of the pollinating insect, bird or other animal, so that pollen is correctly transferred from flower to flower (Goulson 2003). Our native plants have evolved without bumblebees. So feral bumblebees are likely to be ineffective pollinators of some native plants because they do not match the shapes of these native flowers.
For plants that require cross-pollination, the effective pollinator must also carry pollen from one plant to another. Bumblebees tend to take fewer long flights when foraging than other pollinators, which may increase plant self-pollination and reduce seed set (Goulson 2003).
In Tasmania, bumblebees have extensively invaded native vegetation and forage on a very wide range of native plants (Hingston and McQuillan 1998a,b, 1999; Hingston et al. 2002, 2004b, 2006; Hingston 2005a, 2006). The swift parrot has been shown to be ten times more efficient than feral bumblebees in producing seed set in Eucalyptus globulus flowers in Tasmania, probably because bumblebees were self-pollinating this largely self-incompatible native tree (Hingston et al 2004b).
Much more research is needed, but the introduction of feral bumblebees is likely to have a significant effect on the native plants of the mainland. |
|
Bumblebee Impact on Native Birds
|
|
AHGA Report Says
|
|
The Actual Facts
|
| AHGA report page 35
[Re: Hingston and Potts (1998)] Because the swift parrot, Lathamus discolor, is largely dependent on E. globulus pollen and nectar in its summer breeding range in south-eastern Tasmania, the authors raised a red flag, despite the low bumblebee population.
|
|
No statements remotely like this were made in Hingston and Potts (1998). |
|
AHGA report page 35
Hingston and Mallick (2003) again raised the spectre of native birds being negatively impacted upon by competition for nest sites and nectar. Yet competition for nest sites is a non-issue because bumblebees, unlike honeybees, nest primarily underground in old rodent nests.
|
|
Hingston and Mallick (2003) made no mention of competition for nest sites affecting native birds. |
|
AHGA report page 35
[Re: Hingston and Mallick (2003)] Hingston claimed that bumblebees, in concert with honeybees, are threatening the survival of the endangered swift parrot, Lathamus discolor, by feeding on nectar of E. globulus, a remarkable claim not supported by any evidence of any impact.
|
|
In the published Abstract to which the AHGA refers, Hingston and Mallick (2003) state, 'We found that honeybees Apis mellifera and bumblebees Bombus terrestris usually consumed most nectar from these flowers (of Eucalyptus ovata and E. globulus)'.
This evidence clearly supports their conclusion that destruction of foraging and nesting habitat, collisions with human-made structures, and competition for food from introduced honeybees and bumblebees are the principal threats to the swift parrot.
|
| AHGA report page 36
The following information posted by Hingston on Ornithological News and Scuttlebutt (Website of the Echuca and District Branch of Bird Observers Club of Australia (http://users.mcmedia.com.au/ ~stocky/just-heard.html - accessed June 2005), is typical of some of the misinformation being presented to the public...
|
|
Dr Hingston did not post this information on this website and he considers some of the content of the article on the website inaccurate. For example, the statement, 'If Bumble Bees find their way into Victoria's box-ironbark forests, researchers believe it likely that the Swift Parrot will become extinct' cannot be attributed to Dr Hingston because Hingston and Mallick (2003) did not say anything about the effect that the spread of bumblebees into Victoria would have on swift parrots. |
| AHGA report page 36
The Swift Parrot Recovery Plan 2001-2005, put out by the Swift Parrot Recovery Team (2001)
(http://www.deh.gov.au/biodiversity/ threatened/publications/recovery/swift-parrot/) makes no mention of bumblebees or honeybees as any kind of threat to swift parrots.
|
|
This Recovery plan was written in 2001, before Dr Hingston's major research into the impact of bumblebees and honeybees on Swift Parrots was conducted. |
| AHGA report page 36
...in the small northeastern breeding population [of Swift Parrots] located between Launceston and Smithton, where no E. globulus occur, ....
|
|
This AHGA statement is incorrect on three counts. Firstly, the Smithton-Launceston area is in northwestern Tasmania, not the northeast. Secondly, there is no evidence that swift parrots that breed in northwestern Tasmania comprise a separate population. Thirdly, Mallick et al. (2004) estimated that there were between 6,000 and 14,000 mature E. globulus trees cultivated outside commercial plantations between Launceston and Smithton. |
| AHGA report page 37
...bumblebees are not frequent visitors to E. globulus and thus are highly unlikely to compete with swift parrots or any other pollinators...
|
|
Although some studies found that bumblebees were rare visitors to Eucalyptus globulus, others found bumblebees were much more common visitors.
E. globulus was the seventh most frequently visited plant in a survey of native plant use by bumblebees (Hingston and McQuillan 1998a) and, on flowers of E. globulus, bumblebees were between 0.4 and 27.5% of insect visitors in other studies (Hingston and Potts 1998, Hergstrom et al. 2002, Hingston 2002; Hingston et al. 2004a, Hingston et al. 2004b).
|
AHGA report page 59
There is no evidence or logic to the attempt to implicate bumblebees as a threat to native birds feeding on nectar and pollen. It is abundantly clear that man is the key threatening process for all these bird species.
|
|
In Tasmania, bumblebees have extensively invaded native vegetation and forage heavily on flowers that are important food sources for birds (Hingston and McQuillan 1998a; Hingston et al. 2002, 2004b, 2006,). So bumblebees could harm native birds by reducing the amounts of nectar and pollen available to them.
In temperate regions of Australia, nectar surpluses in bird-pollinated plants typically occur in winter and spring, while nectar supply is usually limited in summer and autumn (Paton 2000). However, bumblebee populations also peak in summer and autumn. So the spread of feral bumblebees would put additional pressure on nectar feeding birds during these food-limited seasons.
Whilst it is true that land clearing and habitat fragmentation by man is the key threatening process for many nectar-feeding birds, does man need to make matters worse by also introducing feral bumblebees to the mainland? |
|
Bumblebee Impact on Invasive Weeds
|
|
AHGA Report Says
|
|
The Actual Facts
|
|
AHGA report page 38
As far as we are aware, there has been no impetus for further work to be conducted on the weed issue in Tasmania.
|
|
Two scientists are currently conducting such research. Guy Williams and Dr Andrew Hingston are investigating the effect of bumblebees on seed production in seven species of weeds in Tasmania.
Guy Williams is including research into the effect of B. terrestris on seed production in the weeds tree lupin Lupinus arboreus, English broom Cytisus scoparius and foxglove Digitalis purpurea in Tasmania as part of his PhD at Macquarie University.
Dr Andrew Hingston has produced a list of plants that have become much more invasive in Tasmania since bumblebees arrived, most of which are also invasive in other parts of the world where bumblebees occur, but are not yet serious weeds on the Australian mainland. (Read more)
These are: Rhododendron ponticum (Ericaceae); white-edged nightshade Solanum marginatum (Solanaceae); butterfly bush Buddleja davidii (Loganiaceae); and Agapanthus praecox ssp. orientalis (Liliaceae).
|
AHGA page 54
Published information on 'sleeper' weeds cited as being at risk of spread by B. terrestris gives no credence to the claims that importation of B. terrestris will result in a substantial increase in their spread.
|
|
The AHGA has glossed over this serious weed issue!
Goulson (2003) points out that exotic weeds already cost Australia an estimated AU$3 billion per year in control and lost production. Goulson concludes, 'If even one new major weed occurs in Australia due to the presence of bumblebees, the economic and environmental costs could be substantial'.
Of the 15 potential sleeper weeds nominated by the Weeds Cooperative Research Centre and other specialists (see AHGA report page 51), only three were addressed in any detail by the AHGA and a further five were only briefly mentioned. Two important weeds that the AHGA neglected to discuss are Solanum species and Impatiens.
Solanum -- In Tasmania, the White-edged Nightshade weed, Solanum marginatum, has had its status upgraded from secondary to noxious since feral bumblebees became established. What would happen on the Australian mainland?
Solanum plants require a special kind of pollination called buzz pollination. Honeybees are unable to do this, but bumblebees are buzz pollination specialists. In fact, tomatoes (Solanum lycopersicum) belong to this group and this is a major reason why greenhouse tomato growers wish to import bumblebees for tomato pollination.
Published information (Healy 1976; Roy et al. 1998, Parsons and Cuthbertson 2001) shows that many species of Solanum are weeds of agricultural land and/or disturbed forests. Many produce fleshy fruits readily dispersed by birds and other animals. Livestock have died after eating S. dulcamara or whitehorse nettle S. elaeagnifolium. Moreover, buffalo burr S. rostratum, Jerusalem cherry S. pseudocapsicum and false Jerusalem cherry S. diflorum, are responsible for frequent poisonings of children who eat the attractive fruits, and deaths have resulted.
Impatiens -- Bumblebees could also help Impatiens glandulifera become an invasive weed in Australia. Bumblebees are its major pollinator.
Published information (Clapham et al. 1962; Valentine 1978; Usher 1986; Webb et al. 1988; Sowig 1989; Beerling and Perrins 1993; Perrins et al. 1993; Pysek and Prach 1995; Williamson 1996; Chittka and Schurkens 2001) shows that Impatiens is a weed of river banks, lake sides and other disturbed or damp areas in continental Europe, Britain, North America and New Zealand where it often grows so densely that it smothers other plants. Bumblebees including B. terrestris are its major pollinators.
Over half of the riverbanks in the Czech Republic are currently occupied by Impatiens (Prysek and Prach 1995). However, in the absence of bumblebees on mainland Australia, Impatiens is not an established weed here yet.
|
|
Summary of Bumblebee Impact in Tasmania
|
|
AHGA Report Says
|
|
The Actual Facts
|
| AHGA report page 38
...B. terrestris is located across much of Tasmania, but is rare in areas of native bush, with nests, where present, smaller and less persistent than in urban and semi-urban areas.
|
|
This AHGA statement is totally incorrect.
Bumblebees have proven to be highly invasive in urban, agricultural and native vegetation in Tasmania (Hingston and McQuillan 1998a; Hergstrom et al. 2002; Hingston et al. 2002; Hingston 2006), and are widespread and abundant in native vegetation (Hingston and McQuillan 1998a; Hingston et al. 2002; Hingston 2006) where large and highly successful nests have been observed (Hergstrom et al. 2002; Hingston et al. 2006). One nest examined in a Tasmanian National Park, isolated from urban and agricultural areas by 10km of sea, had produced at least 304 new queens and 939 workers!
Evidence of bumblebee breeding has been found in all of Tasmania's major native vegetation types, from sea level to 1260 m above sea level, across the entire range of annual rainfall (Hingston et al. 2002), and in 10 National Parks and the most remote parts of the Tasmanian Wilderness World Heritage Area (Hingston et al. 2002; Hingston 2006).
|
|
AHGA report page 38
[Re: studies of impact on native bees and native plants] The studies were not definitive because of design flaws, particularly in methodology, and a lack of understanding of bumblebee foraging behaviour and the complexities of this type of study. Pollinator assemblages were broad in the few plants that were studied, and contradictory results were often obtained by Hingston et al. and Hergstrom et al. (2002).
|
|
The AHGA's criticisms of these studies' designs and methodology are largely based on one anonymous 2004 report and a private report commissioned by the AHGA (McClay 2005). Neither of these reports have been presented for examination by readers of the AHGA report; so these criticisms are unproven.
Considering that the work of Hingston et al. on the impacts of bumblebees consists of nine papers published in peer-reviewed scientific journals (Hingston and McQuillan 1998a,b, 1999; Hingston et al. 2002, 2004b, 2006; Hingston 2005a,b, 2006) while the work of Hergstrom et al. (2002) consists of one unreviewed report, the work of Hingston et al. should carry much more weight than does the work of Hergstrom et al. (2002).
|
|
AHGA report page 38
Despite attempts by Hingston to link the demise of threatened bird species such as the swift parrot to bumblebees, there is no evidence to indicate that this is so, quite the contrary. His own studies show limited visitation to E. globulus, a preferred and important nectar resource in swift parrot breeding areas in Tasmania.
|
|
As stated above, E. globulus was the seventh most frequently visited plant in a survey of native plant use by bumblebees (Hingston and McQuillan 1998a) and, on flowers of E. globulus, bumblebees were between 0.4 and 27.5% of insect visitors in other studies (Hingston and Potts 1998, Hergstrom et al. 2002, Hingston 2002; Hingston et al. 2004a, Hingston et al. 2004b). |
|
Would Bumblebees Become Feral on Mainland Australia?
|
|
AHGA Report Says
|
|
The Actual Facts
|
AHGA report page 39
The fact that no establishment has thus far occurred in Australia, despite the more favourable climate on the coast and several attempts at introduction in the past, might be taken as some indication of the difficulties of successful establishment on the mainland. |
|
Feral bumblebees are widespread and abundant in Tasmania and New Zealand, and are also established in Japan, Israel, Chile, Mexico, Argentina and Uruguay. So it is likely that they are capable of also establishing on the Australian mainland.
Unsuccessful attempts in the past to import bumblebees to the Australian mainland could have been due to chance mishaps befalling the small original populations rather than to an unsuitable environment. In fact, the first attempts to establish bumblebees in Tasmania also failed (Council of Agriculture 1909), as did the first attempt to establish honeybees in Australia (Paton 1996). |
| AHGA report page 41
Only large colonies are able to produce reproductive females and males. To achieve this size, a sequence of suitable flowers for 12-15 weeks is needed
|
|
Bumblebees can routinely forage up to 2 km from their nests, and one study predicted that they could forage 10 km from their nests. Therefore each colony normally forages in an area of over 12 square km and might even cover over 300 square km!
Even if one group of suitable plants stopped flowering, the colony would be very likely to find a replacement group of flowers within that area.
|
| AHGA report page 48
Annual rainfall below 400mm will also not support colony development (R. Macfarlane, pers. comm. to S. Goodwin 1997).
|
|
Bumblebees occur in even the driest parts of New Zealand, with mean annual rainfalls as low as 339mm. |
|
Probable Consequences of Bumblebees Becoming Feral
|
|
AHGA Report Says
|
|
The Actual Facts
|
| AHGA report page 50
Across these vast and divergent areas [...] B. terrestris (and all other Bombus species for that matter) is considered only as a benign and efficient pollinator. It has no pest status in any of these areas.
|
|
In fact, bumblebees are considered a threat to the environment by experts in each of the countries and areas below. The threats include transmission of diseases and pathogens, hybridisation with local bumblebee species and competition with native bee species:
Japan
Bombus terrestris is being declared an Invasive Alien Species by the Japanese government.
Importation and use of this bumblebee in greenhouses is only permitted if the greenhouse is fully screened to prevent this bumblebee escaping into the environment.
United Kingdom and Europe
Widmer et al 1998 writes:
In Europe, about 800,000 colonies are currently commercially produced every year for crop pollination in the greenhouse industry[...]. Because they originate from different stocks, commercially bred bumble bees differ genetically from local populations and escaping sexuals may introduce foreign alleles. Therefore, the commercial use of B. terrestris s.l. for pollination should be of concern with respect to genetic changes in local populations.
Chittka et al 2004 writes:
Unfortunately, the diversity of bumblebee populations is currently under threat from indiscriminate import of bee colonies for commercial greenhouse pollination.
Ings et al 2005 writes:
Alarmingly, colonies of these bees have all been shipped in extremely large numbers to regions where other populations are native without appropriate risk assessment. ... new queens (gynes) and males (Goulson et al. 2002a) escape from the glasshouses where they are used as crop pollinators. ... it is imperative for more high quality research to be carried out to provide essential information needed for an overdue risk assessment .. In particular, the risks associated with hybridisation, parasite spread and phenological differences between populations....
Canada
Colla et al 2006 writes:
... pathogens may spread to wild bees when commercial bees escape from greenhouses and interact with their wild counterparts at nearby flowers. ... foraging bumble bees regularly escape from greenhouses during the summer months and up to 73% of the pollen carried by returning workers is collected from plants outside the greenhouse (Whittington et al., 2004). The potential for contact between commercial and wild bumble bees near greenhouses is, therefore, substantial.
United States of America
Thorp 2005 writes:
The major threats to these [North American] bumble bees are exotic diseases introduced via trafficking in commercial bumble bee queens and nests for greenhouse pollination of tomatoes (Thorp 2003, Thorp et al. 2003), habitat loss due to urban development and agricultural conversion, and the extensive use of pesticides.
At least two of the disease organisms [...] are known to occur in commercially produced bumble bee colonies and have been introduced from Europe into Japan (Goka et al. 2000; Goka et al. 2001; Niwa et al. 2004). It is likely that the Nosema outbreak in commercial bumble bee production facilities in North America reported in 1998 (Flanders et al. 2003) is also responsible for the severe declines seen in B. franklini and B. occidentalis [...]. Declines of the other two species of Bombus sensu stricto in eastern North America, B. affinis and B. terricola, (J. Ascher personal communication) probably have the same root cause.
|
| AHGA report page 57
It [Aussie Bee website] also devoted an article to the pleasures of assisting in the export of several nests of an Australian native bee Trigona carbonaria to Japan in 1999 and 2000 (http://www.uq.net.au/~zzrzabel/our-bees-in-japan-2.htm), perhaps suggesting a need to be more objective.
|
|
Firstly, the address of the Aussie Bee website is www.aussiebee.com.au. The article referred to in this AHGA statement actually appears on a completely different website (perhaps suggesting a need for the AHGA authors to be more accurate).
Secondly, introducing the stingless bees, Trigona carbonaria, to Japan is entirely different from introducing bumblebees to Australia for two reasons:
1. Trigona stingless bees are tropical species. The Japanese researcher, Dr Kazuhiro Amano has stated that these bees would not survive the Japanese winter, so they pose no threat to the Japanese environment. The northern limit of the global distribution of stingless bees is in India, some 1,700 km closer to the equator than Japan. In contrast, the AHGA CLIMEX study clearly shows that large areas of Australia have a climate that could allow bumblebees to survive.
2. Trigona stingless bees have a prolonged and painstaking method of nest establishment. Stingless bee workers inside a Japanese greenhouse would need to escape, find a nest site, break back into the greenhouse to return to their hive, then repeat this feat many times a day over several weeks, whilst they gradually provision the new nest site. Only then could a new stingless bee queen move into the new nest site. In contrast, a single fertilised queen bumblebee, escaping from a greenhouse, is capable of establishing a new nest by herself and individual bumblebee colonies can each produce hundreds of new queen bees.
|
| AHGA report page 59
...their [bumblebees'] nesting sites are such that they will rarely be disturbed accidentally.
|
|
Feral bumblebees may nest in old rodent nests (inside sheds or outdoors), compost heaps, piles of leaf litter or grass clippings, woodpiles, old stuffed chairs or sofas, covered drains and cavities underneath concrete paths or houses.
These are all examples of bumblebee nest sites that could easily be disturbed accidentally, especially by a child or family dog.
|
| AHGA report page 59
In another article on native bees (https://www.aussiebee.com.au/faq.html), Aussie Bee reports that 1490 Australian native species can sting, and that they can sting more than once and it is possible to be allergic to the sting. Presumably this is not much of an issue.
|
|
Whilst most Australian native bees can sting, there is no comparison between the impact of a sting from a 22 mm bumblebee queen or 14 mm bumblebee worker, compared with that from a typical native bee. The great majority of our native bee species are less than 10 mm long and most have a sting that is not capable of effectively penetrating the human skin.
Furthermore, almost all of our native bee species are solitary and do not have nest stores of honey or pollen to defend. So they are far more placid than social bees (such as the bumblebee) even if someone is disturbing their nest. In contrast, bumblebees will respond aggressively if their nest is disturbed.
|
|
Commonwealth Government Assessment of Bumblebee Risk
|
|
AHGA Report Says
|
|
The Actual Facts
|
| AHGA report page 62
A nomination ...to have B. terrestris declared a Key Threatening Process under the Environment Protection and Biodiversity Conservation Act 1999...was rejected on the basis that there was insufficient evidence to make such a determination.
|
|
Despite insufficient data having been gathered at the time the nomination was assessed, the Threatened Species Scientific Committee still concluded with this important recommendation:
'The Committee urges that extreme caution be shown in considering any proposal to introduce this species to the mainland. In taking this position, it highlights the concern that many native species are dependent on native pollinators, so it could potentially be a threat in the future.'
|
|
References and Further Reading (Including papers listed in the green column, above.)
Armstrong, J. A. (1979). Biotic pollination mechanisms in the Australian flora - a review. New Zealand Journal of Botany 17, 467-508.
Beekman, M., van Stratum, P., and Veerman, A. (1999). Selection for non-diapause in the bumblebee Bombus terrestris, with notes on the effect of inbreeding. Entomologia Experimentalis et Applicata 93, 69-75.
Beekman, M., and van Stratum, P. (2000). Does the diapause experience of bumblebee queens Bombus terrestris affect colony characteristics? Ecological Entomology 25, 1-6.
Beerling, D. J., and Perrins, J. M. (1993). Impatiens glandulifera Royle (Impatiens royleiWalp.). Journal of Ecology 81, 367-382.
Bell, M. C., Spooner-Hart, R. N., and Haigh, A, M. (2006). Pollination of greenhouse tomaotes by the Australian bluebanded bee Amegilla (Zonamegilla) holmesi (Hymenoptera: Apidae). Journal of Economic Entomology 99: 437-442.
Cardale, J. C. (1993). Hymenoptera: Apoidea. In 'Zoological Catalogue of Australia'. (Eds W. W. K. Houston and G. V. Maynard.) Vol. 10. (CSIRO: Canberra.)
Chittka, L., and Schurkens, S. (2001). Successful invasion of a floral market. Nature 411, 653.
Chittka, L., Ings, T. C., and Raine, N. E. (2004). Chance and adaptation in the evolution of island bumblebee behaviour. Population Ecology 46, 243-251.
Clapham, A. R., Tutin, T. G., and Warburg, E. F. (1962). 'Flora of the British Isles.' (Cambridge University Press: Cambridge.)
Colla, S. R., Otterstatter, M. C., Gegear, R. J., and Thompson, J. D. (2006). Plight of the bumble bee: pathogen spillover from commercial to wild populations. Biological Conservation 129, 461-467.
Council of Agriculture (1909). Bumble-bees. The Agricultural Gazette Tasmania 17, 218, 242.
Donovan, B. J., and Macfarlane, R. P. (1984). Bees and pollination. In 'New Zealand Pest and Beneficial Insects'. (Ed. R. R. Scott.) pp. 247-258. (Lincoln University College of Agriculture: Canterbury.)
Goulson, D. (2003). Effects of introduced bees on native ecosystems. Annual Review of Ecology, Evolution and Systematics 34, 1-26.
Goulson, D., Stout, J. C., and Kells, A. R. (2002). Do exotic bumblebees and honeybees compete with native flower-visiting insects in Tasmania? Journal of Insect Conservation 6, 179-189.
Healy, A. J. (1976). 'Identification of Weeds and Clovers.' (Editorial Services Limited: Wellington.)
Hergstrom, K., Buttermore, R., Seeman, O., and McCorkell, B. (2002). Environmental Research on the Impact of Bumblebees in Australia and Facilitation of National Communication for/against Further Introduction. Horticulture Australia Project No: VG99033.
Hingston, A. B. (1997). The impact of the Large Earth Bumblebee, Bombus terrestris (L.) (Apidae: Apoidea) on Tasmanian ecosystems. Hons Thesis, University of Tasmania, Hobart.
Hingston, A. B. (1998). Temporal and spatial variation in abundances of native bee species on an altitudinal gradient in southern Tasmania. Australian Journal of Zoology 46, 497-507.
Hingston, A. B. (1999). Affinities between southern Tasmanian plants in native bee visitor profiles. Australian Journal of Zoology 47, 361-384.
Hingston, A. B. (2002). Pollination ecology of Eucalyptus globulus ssp. globulus and Eucalyptus nitens (Myrtaceae). Ph.D. Thesis, University of Tasmania, Hobart.
Hingston AB (2005a). Does the introduced bumblebee, Bombus terrestris (Apidae), prefer flowers of introduced or native plants in Australia? Australian Journal of Zoology 53, 29-34.
Hingston, A. B. (2005b). Inbreeding in the introduced bumblebee Bombus terrestris causes uncertainty in predictions of impacts on native ecosystems. Ecological Management and Restoration 6, 151-153.
Hingston, A. B. (2006). Is the exotic bumblebee, Bombus terrestris (Apidae), really invading Tasmanian native vegetation? Journal of Insect Conservation (in press).
Hingston, A. B., and Mallick, S. A. (2003) Are introduced social bees a threat to the endangered Swift Parrot? In 'Proceedings of the Birds Australia Members' Day and Annual General Meeting 2003'. Geography Department, University of Tasmania, 31 May 2003.
Hingston, A. B., and McQuillan, P. B. (1998a). Does the recently introduced bumblebee Bombus terrestris (Apidae) threaten Australian ecosystems? Australian Journal of Ecology 23, 539-549.
Hingston, A. B., and McQuillan, P. B. (1998b). Nectar robbing in Epacris impressa (Epacridaceae) by the recently introduced bumblebee Bombus terrestris (Apidae) in Tasmania. The Victorian Naturalist 115, 116-119.
Hingston, A. B., and McQuillan, P. B. (1999). Displacement of Tasmanian native megachilid bees by the recently introduced bumblebee Bombus terrestris (Linnaeus, 1758) (Hymenoptera: Apidae). Australian Journal of Zoology 47, 59-65.
Hingston, A. B., and McQuillan, P. B. (2000). Are pollination syndromes useful predictors of floral visitors in Tasmania? Austral Ecology 25, 600-609.
Hingston, A. B., and Potts, B. M. (1998). Floral visitors of Eucalyptus globulus subsp. globulus in eastern Tasmania. Tasforests 10, 125-139.
Hingston, A. B., and Potts, B. M. (2005). Pollinator activity can explain variation in outcrossing rates within individual trees. Austral Ecology 30, 319-324.
Hingston, A. B., Marsden-Smedley, J., Driscoll, D. A., Corbett, S., Fenton, J., Anderson, R., Plowman, C., Mowling, F., Jenkin, M., Matsui, K., Bonham, K. J., Ilowski, M., McQuillan, P. B., Yaxley, B., Reid, T., Storey, D., Poole, L., Mallick, S. A., Fitzgerald, N., Kirkpatrick, J. B., Febey, J., Harwood, A. G., Michaels, K. F., Russell, M. J., Black, P. G., Emmerson, L., Visoiu, M., Morgan, J., Breen, S., Gates, S., Bantich, M., and Desmarchelier, J. M. (2002). Extent of invasion of Tasmanian native vegetation by the exotic bumblebee Bombus terrestris (Apoidea: Apidae). Austral Ecology 27, 162-172.
Hingston, A. B., Potts, B. M., and McQuillan, P. B. (2004a). Pollination services provided by various size classes of flower visitors to Eucalyptus globulus ssp. globulus (Myrtaceae). Australian Journal of Botany 52, 353-369.
Hingston, A. B., Potts, B. M., and McQuillan, P. B. (2004b). The swift parrot Lathamus discolor (Psittacidae), social bees (Apidae), and native insects as pollinators of Eucalyptus globulus ssp. globulus (Myrtaceae). Australian Journal of Botany 52, 371-379.
Hingston, A. B., Gartrell, B. D., and Pinchbeck, G. (2004c). How specialized is the plant pollinator association between Eucalyptus globulus ssp. globulus and the swift parrot Lathamus discolor? Austral Ecology 29, 624-630.
Hingston, A. B., Wapstra, H., and Wapstra, A. (2005). 'City meets Bush -- A Guide to Our Local Bushland. Mt Nelson, Tolmans Hill, Sandy Bay.' (Mt Nelson and Lambert Gully Bushcare Groups: Hobart, Tasmania.)
Hingston, A. B., Herrmann, W., and Jordan, G. J. (2006). Reproductive success of a colony of the introduced bumblebee Bombus terrestris (L.) (Hymenoptera: Apidae) in a Tasmanian National Park. Australian Journal of Entomology 45, 137-141.
Hogendoorn, K., Gross, C. L., Sedgley, M., and Keller, M. A. (2006) Increased tomato yield through pollination by native Australian Amegilla chlorocyanea (Hymenoptera: Apidae). Journal of Economic Entomology 99, 828-833.
Ings, T. C., Schikora, J., and Chittka, L. (2005). Bumblebees, humble pollinators or assiduous invaders? A population comparison of foraging performance in Bombus terrestris. Oecologia 144, 508-516.
Macfarlane, R. P., and Gurr, L. (1995). Distribution of bumble bees in New Zealand. New Zealand Entomologist 18, 29-36.
Mallick, S. A., James, D., Brereton, R., and Plowright, S. (2004) Blue-gums Eucalyptus globulus in north-west Tasmania: an important food resource for the endangered Swift Parrot Lathamus discolor. The Victorian Naturalist 121, 36-46.
McClay, A. (2005). Critical review of four studies relevant to the proposed introduction of the European bumblebee Bombus terrestris (L.) into mainland Australia. Report prepared for the Australian Hydroponic and Greenhouse Association. March 2005.
Parsons, W. T., and Cuthbertson, E. G. (2001). 'Noxious Weeds of Australia'. (CSIRO Publishing: Collingwood, Vic.)
Paton, D. C. (1996). 'Overview of feral and managed honeybees in Australia: distribution, abundance, extent of interactions with native biota, evidence of impacts and future research.' (Australian Nature Conservation Agency: Canberra.)
Paton, D. C. (2000). Disruption of bird-plant pollination systems in southern Australia. Conservation Biology 14, 1232-1234.
Perrins, J., Fitter, A., and Williamson, M. (1993). Population biology and rates of invasion of three introduced Impatiens species in the British Isles. Journal of Biogeography 20, 33-44.
Prys-Jones, O. E., and Corbet, S. A. (1991). 'Bumblebees.' (The Richmond Publishing Co. Ltd: Slough.)
Pysek, P., and Prach, K. (1995). Invasion dynamics of Impatiens glandulifera - a century of spreading reconstructed. Biological Conservation 74, 41-48.
Roy, B., Popay, I., Champion, P., James, T., and Rahman, A. (1998). 'An Illustrated Guide to Common Weeds of New Zealand.' (New Zealand Plant Protection Society: Canterbury.)
Sedgley, M., Hogendoorn, K., and Perkins, M. (2004). Domestication of blue-banded bees for greenhouse pollination. Practical Hydroponics and Greenhouses. Jul/Aug Issue 77. pp. 64-65.
Sowig, P. (1989). Effects of flowering plant's patch size on species composition of pollinator communities, foraging strategies, and resource partitioning in bumblebees (Hymenoptera: Apidae). Oecologia 78, 550-558.
Stout, J. C., Kells, A. R., and Goulson, D. (2002). Pollination of the invasive exotic shrub Lupinus arboreus (Fabaceae) by introduced bees in Tasmania. Biological Conservation 106, 425-434.
Thorp, R. W. (2005). Profile: Subgenus Bombus. In Shepherd, M. D., Vaugyan, D. M. and Black, S. H. eds. Red list of pollinator insects of North America. CD-ROM Version 1 (May 2005). The Xerces Society for Invertebrate Conservation. Portland, OR.
Usher, M. B. (1986). Invasibility and wildlife conservation: invasive species on nature reserves. Philosophical Transactions of the Royal Society of London B 314, 695-710.
Valentine, D. H. (1978). The pollination of introduced species, with special reference to the British Isles and the genus Impatiens. In 'The Pollination of Flowers by Insects'. (Ed. A. J. Richards.) pp. 117-123. (Academic Press: London.)
Webb, C. J., Sykes, W. R., and Garnock-Jones, P. J. (1988). 'Flora of New Zealand. Volume IV. Naturalised Pteridophytes, Gymnosperms, Dicotyledons.' (Botany Division, Department of Scientific and Industrial Research: Christchurch.)
Widmer, A., Schmid-Hempel, P., Estoup, A., and Scholl, A. (1998). Population genetic structure and colonization history of Bombus terrestris s.l. (Hymenoptera: Apidae) from the Canary Islands and Madeira. Heredity 81, 563-572.
Williams, J. A., and West, C. J. (2000). Environmental weeds in Australia and New Zealand: issues and approaches to management. Austral Ecology 25, 425-444.
Williamson, M. (1996). 'Biological Invasions.' (Chapman and Hall: London, UK.)
|
|
|
|
|